Inhaltsverzeichnis
WPG1607 Tiere als Vorbilder für Sportarten
| Veranstaltung | Seminar Biomechanik (Justus-Liebig-Universität Gießen) |
|---|---|
| Autoren | Franzi, Jelena, Lisa, Nico |
| Bearbeitungsdauer | 60 Minuten |
| Präsentationstermin | 11.07.2016 |
| Zuletzt geändert | 8.07.2016 |
Einleitung
In diesem Wiki haben wir uns mit der Fragestellung beschäftigt, inwiefern man Tiere und Menschen im Bezug auf den Sport vergleichen kann. Unterschiedliche Tiere verfügen meist über eine bestimmte Art der Fortbewegung. Der Floh beispielsweise bewegt sich durch Sprünge fort, der sich im Wasser befindende Frosch schwimmt. Diese Tiere scheinen Experten für eine charakteristische Bewegungsart zu sein. Wir analysieren, welche besonderen Merkmale sie aufgrund ihrer Spezialisierung aufweisen und vergleichen diese mit denen des Menschen, der sich in einer der Bewegung der Tiere ähnelnden Sportart versucht.
Im ersten Teil des Wikis wenden wir uns der sportlichen Fertigkeit des Squat Jumps zu und vergleichen sie mit den Sprüngen des Flohs. Der zweite Teil beschäftigt sich mit dem Brustschwimmen beim Menschen. Hier stellen wir einen Vergleich zum Frosch an.
Springen
Der Squat Jump beim Menschen
Über die genaue Ausführung des Squat- jumps mit den dazugehörigen Kraft- Zeitverläufen, wurde bereit ein Wiki erstellt. Vergleich hierzu WP1207- Springen. Im folgenden Abschnitt soll daher die Kinetik des Squat- jumps etwas näher beleuchtet werden. Die Grundlage stellt folgende Studie dar:
“Kinematic Analysis of a Squat- jump” von Radenkovic, O. und Stankovic, M. (2012).
Methoden Mit Hilfe oben genannten Studie, sollten qualitative und quantitative Indikatoren des Squat- jumps in den Bereichen Goniometrie und Kinematik erhalten werden. Hierfür führte ein Proband eine Squat- jump aus. Dieser Sprung wurde mit Hilfe von digitalen Videocameras aufgezeichnet und mit einer Software für kinematische Analysen ausgewertet.
Ergebnisse Der Squat- jump wurde in 3 Phasen eingeteilt. A: Rebound- Phase (konzentrische Kraftentwicklung), B: Flugphase und C: Landung (exzentrische Kraftentwicklung).
Der gesamte Sprung dauerte 54 Frames. Er begann mit der Rebound- Phase bei Frame 14, danach folgte die Flugphase. Sie reichte vom 14. bis zum 38. Frame. Die Landung und somit letzte Phase endete mit dem 54. Frame. Die Aufnahmegewindigkeit betrug 1/60 fps (Frames per second). Daraus kann die Dauer der Flugphase mit 0,4 Sekunden errechnet werden.
kinematische Ergebnisse
Während des Sprunges wurden die Geschwindigkeits- und Gleichgewichtsveränderungen der Schulter, der Hüfte, des Knies und des Sprunggelnekes analysiert.
Schulter

In Abbildung 1 ist der Geschwindigkeitsverlauf des Schultergelenkmittelpunktes in der Sagittalbene dargestellt. Die maximale Geschwindigkeit wurde im 12. Frame (roter Strich) erreicht und betrug 3.60 m/s.
Hüfte

In Abbildung 2 ist der Geschwindigkeitsverlauf des Hüftgelenkmittelpunktes in der Sagittalbene dargestellt. Die maximale Geschwindigkeit wurde ebenso im 12. Frame (roter Strich) erreicht und betrug 3.19 m/s.
Knie

In Abbildung 3 ist der Geschwindigkeitsverlauf des Kniegelenkmittelpunktes in der Sagittalbene dargestellt. Die maximale Geschwindigkeit wurde ebenfalls im 12. Frame (roter Strich) erreicht und betrug 3.52 m/s.
Die maximale Geschwindigkeit wurde in allen 3 Gelenken im 12. Frame, der Reboundphase erreicht. Der13.Frame stellt in allen Gelenken den Rebound- Moment dar.
Sprunggelenk
 ]
]
In Abbildung 4 ist der Geschwindigkeitsverlauf des Sprunggelnksmittelpunktes in der Sagittalbene dargestellt. Die maximale Geschwindigkeit wurde hier im 13. Frame (roter Strich) erreicht und betrug 3.14 m/s.
Goniometrische Ergebnisse
Anhand der goniometrischen Messung, sollte die Rotation des Körpers während des Sprunges beurteilt werden.
Hüfte

Abbildung 5 zeigt die relativen Winkelverläufe des Hüftgelenkes, während des gesamten Sprunges. In der Rebound- Phase betrug der Winkel 164° und in der Landephase 170°. Das Maximum wird im 22. Frame (roter Strich) mit 187° erreicht. Es ist zu sehen, dass der Winkel größer wird und von der Flexion in die Retroflexion übergeht. Im der 2. Teil der Flugphase verkleinert sich der Winkel wieder und geht in eine Flexion über.
Kniegelenk

Abbildung 6 zeigt die relativen Winkelverläufe des Kniegelenkes, während des gesamten Sprunges. Das Maximum wird im 14. Frame mit 175° erreicht. Die Winkel reichen in der Flugphase von 160-170°.
Sprunggelenk

Abbildung 7 zeigt die relativen Winkelverläufe des Sprunggelenkes, während des gesamten Sprunges. In der Flugphase betrug der Winkel 30°, 22. Frame (roter Strich). Im Moment der Landung betrug der Winkel des Sprunggelenkes 25°.
In allen drei Gelenken wurde der maximale Winkel nach dem Rebound- Moment erreicht. Danach fallen die Winkel allerdings in allen Gelenken ab.
[Verfasst von Franziska Knaack]

Der Sprung des Flohs
Die Abbildungen und Daten in dem folgenden Wiki-Abschnitt beziehen sich größtenteils auf die Studie von Gregory P. Sutton und Malcolm Burrows (Biomechanics of jumping in the flea, 2010).
Einleitung
Flöhe können Energie in einer federartigen Struktur in den Beinen speichern und wieder freigeben. Eine Theorie ist, dass der Rückstoß der Feder über ein Hebelarmsystem fungiert und so die Tibia und den Tarsus des letzten Beinpaares des Flohs auf den Boden drückt und somit eine größere Abdruckfläche für den Sprung generiert. Des Weiteren ist das Protein Resilin als fast perfekt elastisches Element von Großer Bedeutung für die Sprunghöhe, soll aber in diesem Wiki nicht weiter vertieft werden. Im Folgenden soll erläutert werden, wie die Anatomie des Flohs die Kinetik und Kinematik des Flohs beeinflusst.
Anatomie des Flohs

Der Floh hat 3 Beinpaare, von denen die hintersten am stärksten ausgeprägt sind. Insgesamt besteht zwischen den Beinen ein Verhältnis von 1:1.3:1.9 von vorne nach hinten. Die Hinterbeine haben hierbei eine Länge von 154% der gesamten Körperlänge. Sie rotieren um die Sagittalebene.
Der Thorax geht am hinteren drittel direkt in die Coxa über. Amproximalen Ende der Coxa befindet sich das Resilin Pad, welches bei Kontraktion der Muskeln Energie einspeichert. Am distalen Ende der Coxa ist der Trochanter, der in den Femur übergeht. Ohne ein weiteres Resilin Pad schließt sich distal zum Femur die Tibia und der Tarsus an. Tibia, Femur und Coxa haben in etwa die gleiche Länge. Der Trochanter fällt etwas kleiner und der Tarsus deutlich länger aus. Kurz vor dem Sprung macht sich der Floh ganz klein und baut Spannung auf. Dabei kann er seinen Femur fast parallel zur Coxa schieben, da sich kleine Einkerbungen für den Trochanter an der Rückseite der Coxa befinden, die genau passen.
Der Sprung

Der Sprung an sich ist eine sehr schnelle und synchron ablaufende Bewegung mehrerer Muskeln. In der Initialen Phase ist der Floh zusammengekauert, Femur parallel zur Coxa, Kontraktion der Muskeln und Energieeinspeicherung im Resilin. Der Coxo-Trochanterale Abschnitt des letzen Beinpaares wird ganz nah zum Boden abgesenkt, die Tibia und der Tarsus haben vollständig Bodenkontakt. In dieser Position verharrt der Floh für ca 100ms.
Unmittelbar vor dem Absprung wird der Thorax nach oben rotiert, sodass er sich fast senkrecht zum Boden befindet, die vorderen Beinpaare dienen dabei der Stabilität und haben keinen Bodenkontakt. Die Kontrakturen der Muskeln, die den Femur neben der Coxa halten werden gelöst und somit wird Kraft über die Tibia und den Tarsus auf den Boden übertragen.
Bei dem Take-off wird die Coxa relativ zum Thorax nach dorsal rotiert, der Trochanter wird rapide fast maximal abgesenkt und die Energie im Resilin wird durch das Lösen der Kontrakturen frei. Der Winkel zwischen Tibia und Femur ist 130°, das Gelenk also extendiert und ist mit dem Tarsus die einzige Unterstützungsfläche, die der Floh noch hat. Nach dem Absprung bleiben die Beine in der Streckung und “hängen” unter dem Hauptcorpus.
Kinetik

Zu Beginn der initialen Bewegung des letzten Beinpaares ergibt sich eine Beschleunigung für den Floh von ungefähr 450ms-2. Er erreicht eine Peak Beschleunigung von 1800ms-2und sinkt auf 0ms unmittelbar zum Take-Off ab. Die Folgerung eines solchen Beschleunigungsverlaufs ist eine S-förmige Geschwindigkeitskurve.
Spring-Moment Arm System
Zunächst ist der Moment Arm kurz und es wirkt eine hohe Kraft. Das bedeutet, dass eine kleine Ground Reaction Force (GRF) wirkt. Wenn dann aber das Trochanteral-Coxo Gelenk abgesenkt wird, also noch vor dem Take-Off, dann wird der Moment Arm in sinus manier länger und die Kraft wird geringer. Da aber der arm schneller länger wird, als die Kraft abfallen kann, ergibt sich zunächst ein positives Produkt der beiden Faktoren. Erreicht aber der Moment Arm seine maximale Größe, ist die Kraft noch nicht minimal. Sie sinkt also noch weiter. Ergo ergibt sich ein negatives Produkt. Plottet man das Ergebnis dieses Prozesses, so erhält man eine Kurve, die der Kombination aus elastischen Mechanismen und rotierenden Hebelarmen ähnelt.
[Verfasst von Lisa Langsdorf]
Mensch & Floh
In verschiedenen Sportarten haben sich über die Jahrzehnte hinweg die unterschiedlichsten Sprungtechniken manifestiert. Diese unterschieden sich in ihrer Ausführung enorm. (Vergleiche hierzu WP1207- Springen).
In diesem Abschnitt des Wikis soll nun ein Vergleich zwischen dem Squat Jump des Menschen und dem Flohsprung gezogen werden. Wie bereits in dem ersten Abschnitt erwähnt, kann der Squat Jump in seine 3 Phase eingeteilt werden. In der ersten Phase, der konzentrischen Kraftentwicklung, laufen einige Mechanismen ab, die der Kauerphase des Flohes ähneln. Der Floh schiebt seinen Femur parallel zur Coxa um Spannung aufzubauen, genau wie der Mensch. Es scheint sich also über die Jahrhunderte hinweg als effizient erwiesen zu haben, aus einer möglichst niedrigen Position nach oben zu springen. Es liegt nahe, dass diese Tatsache der Theorie des Feder-Masse-Modells zugrunde liegt (vergleiche hierzu LM1 -Laufmodelle).
Wenn man nun einen Bezug zwischen dem Feder-Masse-Modell und dem Sprung herleiten möchte, kann man die konzentrische Phase des Sprungs als Kompression der Feder sowie die exzentrische Phase des Sprungs als Dekompression der Feder bezeichnen. Analog kann dann erörtert werden, dass durch die konzentrische Phase des Sprungs (Kompression der Feder) potentielle Energie eingespeichert wird und bei der Exzentrik (Dekompression der Feder) zur maximalen Kraftentfaltung beiträgt. Es scheint eine Analogie dieser Theorie in der Mensch und in der Tierwelt zu bestehen.
Um die Möglichkeit zu haben, sich möglichst lange und somit möglichst effektiv vom Boden abzudrücken, drücken wir Menschen uns mit den Fußspitzen ab. Da der Floh eine andere Beinanatomie hat, ist der Vorgang zwar nicht analog, aber nach dem gleichen Prinzip geschaltet. Sein hinterstes Beinpaar setz so auf dem Boden auf, dass das was bei uns das Sprunggelenk wäre, ventral ist und der Fuß nach dorsal ausläuft. Sein Fuß ist quasi um 180° gedreht im Vergleich zum menschlichen Fuß. Aber auch hier lässt sich vermerken, dass der Floh sich möglichst lange vom Boden abdrückt.
Ein Parameter, der die Sprunghöhe beeinflusst, ist die isometrische Kraft der Beinmuskulatur. Je größer diese wirkende Kraft, desto höher ist die Sprunghöhe. Bei dem Menschen ist der Unterschied zwischen einem Countermovement Jump (Vergleiche hierzu WP1207- Springen) und einem Sprung mittels Katapult Mechanismus kaum unterschiedlich, weil die Bodenreaktionskraft (BRK) während des Absprungs nur ein kleines Vielfaches des eigenen Körpergewichtes ist (2 bis 3 * BW). Der Floh hingegen bringt das bis zu 135-fache seines eigenen Körpergewichtes als BRK zustande. Deswegen macht es für ihn mehr Sinn, einen „Katapultsprung“ zu machen. Unter dem Katapultsprung ist zu verstehen, dass der Floh, wie viele weitere Insekten elastische Energie speichern und sehr rapide verwerten können, ähnlich des DVZ (WP1208 Titin). Da der Floh allerdings das so hohe Kräfte aufbringen muss, um hoch zu springen, müssen sich seine Muskeln behelfen. Der Katapult Mechanismus befähigt die Muskeln, langsam verrichtete Arbeit zu speichern und in eine schnell auslösenden Antwort umzuwandeln. Das begünstigt den Absprung und die ungemeinen Kräfte, die der Floh aufbringt.
Da der Mensch allerdings solche Kräfte nicht aufbringen muss, hat die Natur diese physiologischen Vorteile auch nicht vorgesehen. Der Mensch kann seine Sprunghöhe allerdings durch den DVZ während eines Countermovement Jumps erhöhen.
[Verfasst von Lisa & Franziska]
Schwimmen
Das Bewegen im Wasser stellt für den Menschen, nach der für ihn natürlichen Bewegung auf dem Land, ein weiteres Medium dar. Der folgende Abschnitt beschäftigt sich mit der Analyse des Bewegungsablaufes im Brustschwimmen beim Menschen und soll später mit der zunächst ähnlich erscheinenden Schwimmbewegung des Frosches verglichen werden. Dabei wird genauer auf die verschiedenen Möglichkeiten zur Anpassung an unterschiedliche Techniken eingegangen. Hier gibt es sowohl bei Menschen als auch beim Frosch interessante Parallelen und Unterschiede.
Die Geschichte des Schwimmens, sowie biomechanische Grundprinzipien und Wettkampfregeln können dem Wikieintrag WP1404 – Schwimmen entnommen werden. Für das Verständnis der später folgenden Funktionsanalyse müssen zunächst einige zusätzliche Begriffe geklärt werden
Gesetze für die Fortbewegung im Wasser
Wasser hat eine Dichte von genau 1, was bedeutet, dass die Moleküle gegeneinander verschiebbar sind, wodurch die Gegenkraft des Wassers selbst verringert wird. Das Vorwärtskommen wird jedoch durch die hohe Dichte erschwert. Weiterhin verfügt Wasser aufgrund seiner höheren Dichte im Vergleich zur Luft über einen höheren Widerstand, wodurch der sich im Wasser befindliche Körper abgebremst wird. Ob ein Körper im Wasser schwimmt oder sinkt, hängt von seiner eigenen Dichte ab. Da die Dichte des menschlichen Körpers um den Wert 1 schwankt, können manche Menschen mühelos auf dem Wasser liegen, während andere Menschen, welche einen höheren Anteil an Muskelmasse besitzen, Schwierigkeiten damit haben. (Vgl. Wick, 2013)
Für eine selbsterzeugte Fortbewegung im Wasser wird stets eine wechselseitige Wirkung von Körper- und Wasserbewegung benötigt. Durch einen Kraftstoß des Körpers kommt es zu einer Impulsänderung des Wassers. Dadurch kann der Körper dann schlussendlich fortbewegt werden. Diese wechselseitige Rückwirkung nennt man auch Widerlager. Beim Schwimmen ist das Widerlager abhängig von der Strömung und der unterschiedlichen Drücke um die Antriebsflächen. Einen weiteren wichtigen Faktor, um sich im Wasser vorwärts bewegen zu können, stellt der Sog dar, auf den hier allerdings nicht genauer eingegangen werden soll. Des Weiteren kommt es zu einer Rotation der Wassermassen hinter einem sich bewegenden Körper. Eine geordnete Rotation bezeichnet man als Vortex. Dieses Phänomen kann außerdem zum Antrieb genutzt werden. (Vgl. Ungerechts et al., 2002)
Im Sportschwimmen steht natürlich vor allem die Schnelligkeit im Vordergrund. Um im Brustschwimmen die geforderte Distanz in möglichst kurzer Zeit zurück zu legen, ist es von elementarer Wichtigkeit, eine Technik zu finden, die den Geschwindigkeitsabfall innerhalb eines Schwimmzyklus möglichst gering hält (Vgl. Kandolf, 2011). Laut einer Studie von Yeater et al. (1981) scheint es eine optimale Schlagrate (stroke rate) beim Kraulschwimmen zu geben. An diesem Punkt liegt die Geschwindigkeit bei 0,8 - 0,9 m/s. Nach Überschreiten dieser optimalen Rate scheinen Kraft und Geschwindigkeit wieder abzunehmen. Es bleibt in der Studie noch unklar, ob sich dieser Effekt beim Brustschwimmen ebenfalls in abgeschwächter Form zeigen könnte. Argumente für das Ausbleiben dieses Effekts sind jedoch eine verminderte Armgeschwindigkeit und ein einfacheres Koordinationsmuster.
Brustschwimmen

{kind=link}
Das Brustschwimmen ist eine sehr verbreitete und zudem auch eine der ältesten Schwimmtechniken. Im Sportschwimmen gilt sie als langsamste Art, um sich im Wasser fortzubewegen. Der Antrieb erfolgt durch eine symmetrische, aber phasenverschobene Arm- und Beinaktion. Man unterscheidet zwischen folgenden drei Techniken:
- Undulationstechnik
- Überlappungstechnik
- Gleittechnik
Die Undulationstechnik wird vor allem im Wettkampfschwimmen angewendet und bedient sich einer leicht wellenförmigen Bewegung des Körpers. Die Gleittechnik ist motorisch nicht ganz so anspruchsvoll und findet daher oft im Nachwuchssport Anwendung. Der Körper wird hierbei nicht ganz so weit aus dem Wasser gehoben wie bei der Undulationstechnik und die wellenförmige Bewegung entfällt. Zudem kommt es hier im Gegensatz zum Schwimmen in der Gleittechnik zu keiner Gleitphase. Die Gleittechnik wird vor allem im Freizeit- und Breitensport verwendet. Sie zeichnet sich durch eine wechselnde Antriebsaktion der Arme und Beine aus. Zudem kommt es zu einer Gleitphase, die eine kurze Entspannung nach jedem Zyklus ermöglicht. (Vgl. Ungerechts et al., 2002)
Im Folgenden beschäftigen wir uns mit der Gleittechnik, da diese am besten geeignet ist, um sie später mit der Schwimmbewegung des Frosches zu vergleichen. Die Gleittechnik zeichnet sich durch eine Pause nach der Arm- und Beinbewegung aus. Dies ermöglicht dem Schwimmer, für kurze Zeit aktionslos durchs Wasser zu gleiten. Die folgende Aktionsskizze stellt den Bewegungsablauf nach Ungerechts et. al (2002) stichpunktartig dar.
| Tabelle 1: Aktionsskizze des Brustschwimmens (Ungerechts et. al, 2002, S. 89) |
| Phasen des Brustschwimmens |
|---|
| Arme öffnen und einwärts bewegen |
| Arme vorbringen und strecken |
| Unterschenkel anschwingen |
| Beine kreisförmig strecken |
| Einatmen am Ende der Einwärtsbewegung der Arme, davor ausatmen |
Um den Ablauf besser nachvollziehen zu können, ist hier die Bewegung nochmal visuell veranschaulicht:
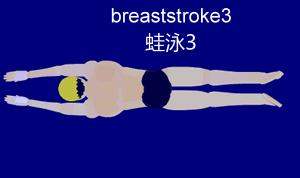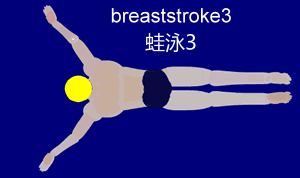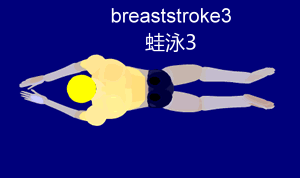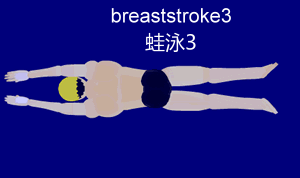
{kind=link}
[Verfasst von Jelena Müller]
Frosch
Basierend auf zwei Studien ( „Built for rowing: frog muscle is tuned to limb morphology to power swimming“ und „Two distinct gait types in swimming frogs“ ) wird zunächst gezeigt, auf welche Probleme der Frosch stößt, wenn er sich im Wasser bewegen will. Danach wird die Frage, wann ein Frosch nicht beidbeinig-synchron schwimmt, beantwortet.
Wie der Frosch schneller schwimmen kann
Frösche sind ein ideales Model um die Grenzen des Muskel-angetriebenen Schwimmens zu untersuchen. Wenn ein Frosch schneller schwimmen will, müsste dieser seine Muskeln schneller verkürzen. Wenn jedoch die Muskeln schneller verkürzt werden, dann fällt die Maximalkraft. Außerdem steigt der Wasserwiderstand mit Zunahme der Geschwindigkeit. Was das genau bedeutet und wie es möglich ist, dass Frösche dennoch schneller schwimmen können, wird im Folgenden erläutert.
Grenzen einer schnelleren Fortbewegung im Wasser
Schauen wir uns zunächst die Abbildung Frosch-1 an. Die Kraft-Geschwindigkeits-Kurve („Muskel F-V Kurve“) (siehe auch Wikieintrag: Kraft-Geschwindigkeits-Verhältnis) zeigt, dass bei geringer Geschwindigkeit der Muskelkontraktion ($V_{Muskelkontraktion}$) hohe Maximalkräfte vom Muskel erzeugt werden können. Nehmen die Muskelkontraktionen in der Frequenz zu, so fällt die nutzbare Maximalkraft („Muskel F-V Kurve“) monoton bis auf 0 ab. Die Kurve zeigt uns demnach das Maximum der Kraft, welche verfügbar ist. Je schneller der Frosch seine Muskeln kontrahiert, desto ineffizienter ist sein schwimmender Vorwärtsantrieb.
Darüber hinaus wird der Muskelarbeitsbereich durch den Wasserwiderstand einschränkt. In der Flüssigkeitsdynamik (siehe Wikieintrag: Strömungsdynamik) wird beschrieben, dass der Wasserwiderstand direkt proportional zum Quadrat der Geschwindigkeit ist (F~v²) (siehe Abb. Frosch-1: „Strömungswiderstand - V Kurve“). Das heißt sein Wirkbereich liegt oberhalb der letzteren Kurve und unterhalb von der „Muskel F-V Kurve“. Einfacher ausgedrückt, damit der Frosch im Wasser (schneller) schwimmen kann, muss dieser den Wasserwiderstand überwinden.
Die Abbildung Frosch-1 stellt dar, dass der Arbeitsbereich des Muskels unterhalb der Kraft-Geschwindigkeits-Kurve und oberhalb der Strömungswiderstand-Kurve liegt (hellgrauer Bereich).
Weiterhin ist in der Abbildung die Überschneidung der beiden Funktionen zu sehen. Sie zeigt die maximal mögliche Geschwindigkeit ($V_{max; morph}$) bedingt durch den Bau des Frosches an.

Möglichkeiten des Froschs um schneller zu schwimmen
Um schneller zu schwimmen müsste eine der beiden Kurven in der Abbildung „angehoben“ werden. Erste Variante, die Kraft-Geschwindigkeits-Kurve könnte „angehoben“ werden (Abb. Frosch-1: Verschiebung von der Strich-Strich-Linie zur schwarzen Linie), indem der Frosch einen neuen Muskel erhält, der mehr Kraft erzeugen kann. Diese naturgegebene physiologische Grenze kann der Frosch nicht ändern, außer er tauscht seinen Körper mit dem eines Froschs mit größeren Muskeln.

Zweite Variante, der Frosch ändert seine Morphologie, also seine Anatomie, um die Kraft besser auf seine Umgebung zu übertragen. Dazu hat er zwei Möglichkeiten. Einerseits kann er die Oberfläche seiner Füße vergrößern (siehe Abbildung Frosch-3 (a-c) (hellblaue Fläche)) und somit die Kraft effizienter auf das Wasser übertragen oder andererseits schaltet er wie ein Auto in den nächsthöheren Gang und schwimmt somit schneller. In der Abbildung „Frosch-2“ sehen wir, wie sich dann die Funktionskurve verändert und der Schnittpunkt (welcher $V_{max;morph}$ darstellt) nach links wandert.
Gänge beim Frosch
In der Abbildung Frosch-3 (a) und (b) sehen wir, wie der Muskel plantaris longus den Winkel $\theta$ verändern kann. Diese Veränderung der Hebelverhältnisse R/r stellt einen Wechsel in einen anderen Gang dar mit welchem sich der Frosch schneller (bzw. langsamer) fortbewegen kann (Gang-Verhältnis = R/r). Außerdem könnte der Muskel (rot) durch eine größere Füße (hellblau) mehr Kraft auf das Wasser übertragen.
(Abb. ist hinter diesem Link: "Figure 3 a" )
(Abb. Frosch-3)
An diesem Frosch-Modell sind die Hebelverhältnisse R/r und der Winkel $\theta$ dargestellt. Sie haben Einfluss auf die Geschwindigkeit.
Erklärungen des Frosch-basierten Muskel-Modelles
Das Modell kann die Performance ($V_{max}$) verschiedener Frösche beim Schwimmen erklären in Abhängigkeit von ihrer jeweiligen Morphologie (Gänge, Größe der Füße) und ihrer Physiologie der Muskeln (Durchmesser und somit $V_{max}$, $F_{max}$). Wichtig ist demnach, dass auf die Morphologie des Körpers und die Muskeleigenschaften geschaut werden muss, um Vorhersagen für die Maximal Geschwindigkeit zu machen. Die Studie zeigt auch, dass es nicht eine einzige optimale Morphologie gibt um sich fortzubewegen. Deshalb kann dieses Modell eventuell genutzt werden um unter evolutionären Gesichtspunkten zu erklären warum einige Frösche größere Füße, ein anderes Kraft-Umsetzungsverhältnis in den Beinen oder andere kräftigere Muskeln haben.
[Verfasst von Nico]
Wann der Frosch mit einer ungleichmäßigen Beinbewegung schwimmt
Wenn eine Gangart zu energie-intensiv wird, dann wechseln Tiere meistens ihre Gangart in eine weniger energie-intensive. Wenn wir zum Beispiel beim Laufen immer langsamer werden, beginnen wir ab einem bestimmten Punkt mit dem Gehen. Genauso verhält es sich im Tierreich. Oben im Text war zu sehen, dass der Frosch auch verschiedene Gänge hat. Dies bezog sich auf phasengleichförmige synchrone Beinbewegungen.
Daneben gibt es noch eine andere Technik für die langsame Vorwärtsbewegung, die „phasenverschobene“ Beinbewegung. Hierbei bewegen sich die Beine nicht synchron, also asynchron (siehe Video: „Frosch schwimmt phasenverschoben“). Zunächst wurde davon ausgegangen, dass die phasenverschobene Beinbewegung nur selten und für Richtungsänderung benutzt wird. Wissenschaftler (S. Nauwelaerts und P. Aerts) haben beobachtet, dass diese Art der Fortbewegung sehr oft genutzt wird. Außerdem haben sie berechnet, dass bei gleicher Geschwindigkeit, die phasenverschobene Beinbewegung weniger Energie erfordert, als die gleichphasige. Sie erklären das wie folgt: Während des phasenverschobenen Schwimmens variiert die Geschwindigkeit des Frosches weniger, weil sich das linke und rechte Bein abwechseln um Vorwärtsantrieb zu erzeugen. Wie bereits erwähnt, gilt für den Wasserwiderstand: F~v². Somit erhöhen stärkere Geschwindigkeitsänderungen den mittleren Wasserwiderstand.
Frosch schwimmt phasenverschoben
Link zu youtube: Frosch schwimmt phasenverschoben
[Verfasst von Nico]
Mensch und Frosch
Dass es sich beim Schwimmen um einen Techniksport handelt, ist allseits bekannt und wurde bereits zuvor erwähnt. Um seine Technik zu variieren hat der Mensch die Möglichkeit, zwischen unterschiedlichen Stilen (Brust, Rücken, Kraul, Delfin) zu wechseln. Innerhalb dieser Stile, wie beim Brustschwimmen, können weitere Technikvariationen auftreten (Überlappungs-, Gleit-, Undulationstechnik). Der Frosch kann seinen Schwimmstil nicht wechseln.
Betrachten wir bei beiden das Brustschwimmen, so können beide durch eine Veränderung ihrer Koordination und/ oder zeitliche Abfolge sowie Stellung der Gliedmaßen die Technik variieren. Im Gegensatz zum Menschen bewegt der Frosch seine Beine auch phasenverschoben beim schwimmen, also nicht gleichzeitig synchron. Der Frosch benutzt diese Bewegungstechnik auch um die Richtung zu wechseln, während der Mensch eher die Arme dazu nutzt.
Außerdem ist der Frosch in der Lage aufgrund seiner anatomischen Besonderheiten innerhalb seiner Brustschwimmtechnik zu variieren. Der Frosch macht sich seine Anatomie zunutze, indem er die Fläche seiner Hände, aber vor allem seiner Füße, durch seine Schwimmhäute variieren kann und durch verschiedene Beingelenkstellungen, den Gängen, die Kraftübertragung auf das Wasser beeinflussen, kann. Im Gegensatz dazu kann der Mensch auch mit technischen Hilfsmitteln seine „Anatomie“ beeinflussen indem er Hand- oder Fußflossen benutzt oder einen Schwimmanzug anzieht um den Reibungswiderstand zu verringern (siehe Wiki zum Thema Reibung)
Ein technischer Unterschied zwischen Frosch und Mensch ist, dass der Frosch die Kraft für die Fortbewegung fast ausschließlich nur aus den Beinen gewinnt und der Mensch die Arme mehr als die Beine nutzt.
Wie bereits erwähnt, spielt beim Menschen das Sportschwimmen eine große Rolle. Hier ist das Ziel eine relativ kurze Strecke möglichst schnell zu überwinden. Die schnellsten Zeiten werden dabei im Delfin- und Kraulstil erzielt. Das Brustschwimmen gilt als eher langsame Art der Fortbewegung im Wasser. Daher vermuten wir, dass dieser Stil besonders für längere Strecken gut geeignet ist. Hier könnte sich eine weitere Parallele zum Frosch auftun. Dieser schwimmt möglicherweise im Bruststil, da er nicht unbedingt schnell sein muss, sondern eine für ihn weite Distanz überwinden muss und dies durch den Bruststil am ökonomischsten ausführen kann.
[Verfasst von Jelena & Nico]
Zusammenfassung und Ausblick
Für die Zukunft wäre die Verbindung zwischen anderen Sportarten und Tieren interessant. Der Forschungsstand bezüglich der Kinetik von tierischen Bewegungen im Bezug auf den Menschen sind allerdings noch nicht sehr ausgeprägt und erfordern noch wissenschaftliche Untersuchung. Betrachtet man allerdings den Fachbereich der Materialwissenschaft, ist zu vermerken, dass die Forschung schon deutlich weiter ist, als die Sportwissenschaft. Im Rahmen der Bionik wurden schon beträchtliche Ergebnisse gewonnen.
Fragen
Fragen für den Floh
Was ist ein großer Bestandteil der Kraftentfaltung in der Exzentrik? <spoiler | Antwort> Die konzentrische Kraftentfaltung. Hier wird potentielle Energie eingespeichert und in der Exzentrik gelöst. </spoiler> Was sind die drei Phase des Squat Jumps? <spoiler | Antwort> Rebound-Phase, Flugphase, Landung. </spoiler> Welche Art der Kraftentwicklung findet in diesen Phasen statt? <spoiler | Antwort> Rebound-Phase: konzentrische Kraftentwicklung.Die Flugphase ist isoton. Landung: Exzentrische Kraftentwicklung. </spoiler> Was ist die Besonderheit der Beinanatomie des Flohs? <spoiler | Antwort> Er kann seinen Femur parallel neben die Coxa flektieren. Außerdem ist sein „Fuß“ um 180° gedreht. </spoiler> Welches Modell liegt der Aussage zugrunde, dass man aus einer möglichst niedrigen initialen Position starten sollte, um hoch zu springen? Erläutere dieses kurz. <spoiler | Antwort> Feder-Masse-Modell. In der konzentrischen Phase wird potentielle Energie gespeichert. Das geht mit einer maximalen Kraftentfaltung in der exzentrischen Phase einher. Daraus folgend wird eine möglichst große Sprunghöhe erzielt </spoiler>
Fragen für Frosch
Erkläre synchrones und asynchrones schwimmen beim Frosch? Vor-und Nachteile? (Stichwort: beidbeinig; Wasserwiderstand) <spoiler | Antwort> Meistens schwimmt der Frosch beidbeinig synchron, das eignet sich gut für das schnelle Schwimmen. Das asynchrone Schwimmen (beide Beine phasenverschoben) eignet sich eher für langsame Geschwindigkeiten. Außerdem gilt für den Wasserwiderstand: F~v². Werden die Beine abwechselnd (asynchron) bewegt, ist die Kraftzufuhr für die Vorwärtsbewegung kontinuierlicher. Daraus folgt, dass der mittlere Wasserwiderstand der überwunden werden muss geringer ist. Das asynchrone Schwimmen ist folglich kräftesparend. </spoiler>
Was könnte der Frosch tun, damit er schneller schwimmt? (erkläre die Kurven in Grafik) <spoiler | Antwort> Um schneller zu schwimmen muss der Frosch einen größeren Wasserwiderstand überwinden, dazu muss er mehr Kraft erzeugen. Entweder er macht Krafttraining oder tauscht seine Muskeln aus. Eine andere Möglichkeit ist, dass er seine Morphologie ändern kann. Das heißt einerseits die Oberfläche der Füße vergrößern oder andererseits in den nächsten Gang schalten (Gelenkswinkel ändern) um eine bessere Kraftübertragung zu erreichen.</spoiler>
Welche Probleme treten auf, wenn der Frosch schneller schwimmen will? <spoiler | Antwort> Er müsste die Muskeln schneller verkürzen (kontrahieren). Das Problem ist, dass damit die Maximalkraft abnimmt (physiologische Grenze). Weiterhin ist problematisch, dass der Wasserwiderstand bei größer Geschwindigkeit zunimmt, daraus folgt, dass der Frosch mehr Kraft benötigt um schneller zu schwimmen ( F~v²). </spoiler>
Was ist der Unterschied zwischen $v_{max}$ und $v_{max; morph}$? <spoiler | Antwort>
- $v_{max}$: Geschwindigkeit, die der Muskel erzeugen kann (ohne Berücksichtigung von Wasserwiderstand)
- $v_{max; morph}$: Geschwindigkeit, die von Morphologie / Anatomie abhängt, d.h. Stellung der Beine (Gelenkswinkel in der Hüfte, Knie, Fußgelenk)
</spoiler>
Literatur
Alexander R. McN., (1995), Leg Design and Jumping Technique for Humans, other Vertebrates and Insects
Radenkovic O., Stankovic M. (2012), Kinematic Analysis of a Squat-Jump
Sutton G.P., Burrows M. (2010), Research Article Biomechanics of jumping in the flea
Rothschild M. et al (1972), Jump of the Oriental Rat Flea Xenopsylla cheopis
Kandolf W., Spezielle Biomechanik und Bewegungslehre sowie Trainingslehre des Schwimmens, IFS Universität Innsbruck (2011)
Nauwelaerts S. , Aerts P. (2002), Two distinct gait types in swimming frogs: Journal of Zoology, Vol 258, Issue 2, S. 183 – 188.
Richards C. T., Clemente C. J. ( 2013). Built for rowing: frog muscle is tuned to limb morphology to power swimming: Journal of the Royal Society Interface
Shimojo H., Sengoku, Y., Miyoshi, T., Tsubakimoto, S., Takagi, H. (2014). Effect of imposing changes in kick frequency on kinematics during undulatory underwater swimming at maximal effort in male swimmers. Human Movement Science 38 (2014), S. 94 - 105.
Ungerechts B., Volck, G., Freitag, W. (2002). Lehrplan Schwimmsport - Band 1: Technik. Schorndrof: Verlag Karl Hofmann
Wick D. (2013). Biomechanik im Sport - Lehrbuch der biomechanischen Grundlagen sportlicher Bewegungen.(3., überarbeitete und erweiterte Auflage). Balingen: Spitta Verlag GmbH & Co. KG
Yeater R. A., Martin R. B., White M. K., Gilson K. H. (1981). Tethered Swimming Forces in the Crawl, Breast and Back Strokes and their Relationship to Competitive Performance. J. Biomechanics, 14 (8), S.527 - 537.
Bewertung des Wiki-Moduls
| Kategorie | Punkte | Anmerkungen |
|---|---|---|
| Inhalt (max. 10) | 9 Pkt | knappe Einleitung (wenig beispielhaft; nicht durch Medien bestärkt), ebenso knapper Ausblick/Fazit, kritische Betrachtung wünschenswert |
| Form (max. 5) | 4 Pkt | am Anfang wenig Verlinkungen, einige Grafiken zu kleine Beschriftungen |
| Bonus (max. 2) | 0 Pkt | |
| Summe | 13/15 Pkt | 86,7 % |